
 A fejlődés során becsúszó mutációk, az epigenetikai szabályozás és a környezeti hatások miatt, nincs két tökéletesen egyforma élőlény, bár értelemszerűen a klónok, partenogenetikus egyedek és az egypetéjű ikrek meglehetősen közel állnak ehhez. A genetikai anyag közel tökéletes megegyezése az, ami igazán különbözővé teszi pl. utóbbiakat a kétpetéjű ikrektől, amelyek csak éppen annyira hasonlítanak mint egy átlagos testvérpár (hiszen szintén két különböző petesejt és spermium egyesülésével jönnek létre).
A fejlődés során becsúszó mutációk, az epigenetikai szabályozás és a környezeti hatások miatt, nincs két tökéletesen egyforma élőlény, bár értelemszerűen a klónok, partenogenetikus egyedek és az egypetéjű ikrek meglehetősen közel állnak ehhez. A genetikai anyag közel tökéletes megegyezése az, ami igazán különbözővé teszi pl. utóbbiakat a kétpetéjű ikrektől, amelyek csak éppen annyira hasonlítanak mint egy átlagos testvérpár (hiszen szintén két különböző petesejt és spermium egyesülésével jönnek létre).
A kétfajta iker-rokonság közötti átmenet kifejezetten nehezen elképzelhető, ha nem éppen lehetetlenül hangzik, különösen emberek esetén (bár.... - lásd kicsit lejjebb), de más fajok nem ennyire fantáziátlanok.
Példának okáért ott vannak a selyemmajmocskák, ahol már hosszabb ideje ismert, hogy a méhben fejlődő (kétpetéjű)ikrek a fejlődés során szorgalmasan cserélgetik egymás között vérképző őssejtjeiket, így például az sem ritka, hogy kifejlett hímekben XX kromoszómás vérsejtekre leljünk, amelyek a nővérétől még a méhben kapott ősvérsejtekből keletkeznek (illetve, természetesen, vice versa). Arra azonban, csak most derült fény, hogy ez a fajta csere-bere nem szorítkozik a vérképző őssejtekre, hanem gyakorlatilag bármilyen őssejt alanya lehet. És a "bármilyen" az az ősivarsejtekre is vonatkozik. Bizony, akármilyen hihetetlenül hangzik, a selyemmajmok között nem ritka, hogy a büszkén párzó hím éppen nem saját genetikai állományának örökítésén mesterkedik, hanem ikerbátyja DNS-ét adja tovább és szexuális vonzalmának tárgya sem (genetikailag) saját petesejtjét kínálja fel megtermékenyítésre, hanem nővéréjét. És mivel akár a fiú testvér ősivarsejtje is átkerülhet egy-egy nősténybe, az a bizarr helyzet is előállhat, hogy a petesejt Y kromoszómát tartalmaz! (Egyébként ez a megfigyelés "mellékesen" azt is bizonyította, hogy megfelelő molekuláris környezetben a hím-ősivarsejtek in vivo is képesek petesejtet produkálni. Ennek a kísérletnek az in vitro megfelelőjét alig pár hónapja végezték el, egerekben.)
Persze maga a megfigyelés egy elég nehéz kérdést vet fel: miért csinálják ezt a selyemmajmok? Jelenleg csak egy kicsit erőltetett magyarázat létezik, miszerint az egész arra vezethető vissza, hogy a egyes fajok nőstényei több hímmel is párzanak és ha a utódaik ilyen kimérák lesznek, akkor minden apa a sajátjaként fogja kezelni őket (ebben a felismerésben fontos szerepet játszanak a bőrből származó feromonok). Ezt látszik alátámasztani az a megfigyelés is, hogy a kiméra-bébikkel sokkal többet foglalkoztak az apák, mint a nem kimérákkal, de hogy az anyai gondoskodás miért mutat pont fordított korrleációt, az mindenképpen misztikus.
 Ha pedig már különleges ikreknél tartunk, akkor ez remek apopó, hogy egy igen különleges emberi példáról is írjak. Olyan ikerpárról van szó, amelyik egyik tagja kisfiú, a másik .. nos .. az pedig hermafordita: hím és női ivarszervei egyaránt vannak. Ezen kívül ún. ovotestisük van, amelyben egymás közelében fejlődnek a hím és női ivarsejtek.
Ha pedig már különleges ikreknél tartunk, akkor ez remek apopó, hogy egy igen különleges emberi példáról is írjak. Olyan ikerpárról van szó, amelyik egyik tagja kisfiú, a másik .. nos .. az pedig hermafordita: hím és női ivarszervei egyaránt vannak. Ezen kívül ún. ovotestisük van, amelyben egymás közelében fejlődnek a hím és női ivarsejtek.
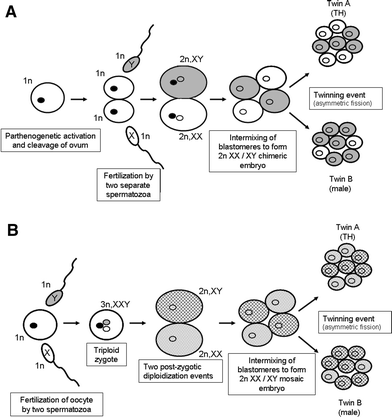
A hermafroditizmus önmagában is ritkaságszámba megy, de ami itt igazán felkeltette az orvosok figyelmét az az volt, hogy a kisbaba egyes sejtjei XX, mások pedig XY kromoszómájúak voltak. Közelebbi vizsgálat kiderítette, hogy testvére esetében is előfordul mindkét sejttípus, csak ott az XX sejtek aránya kisebb, ezért fejlődhetett gond nélkül kisfiúvá. Pusztán ezek alapján az tűnne a legkézenfekvőbb magyarázatnak, hogy egy kétpetéjű ikerpár sejtjei valami különös véletlen folytán összekeveredtek. A valóság azonban rácáfolt erre: ami leginkább megdöbbentette a dokikat az a genetikai markerek vizsgálata volt: ezek ugyanis azt mutatták, hogy a gyerekek egyetlen petesejt, és két különböző (bár egy apától származó) spermium találkozásából jöttek létre. Kvázi másfélpetéjű ikrekről van tehát szó.
Egy ilyen édes-hármas találkozásnak ismereteink szerint igen kicsi a valószínűsége, annak pedig mégkisebb, hogy két életképes kisbabát eredményezzen. Így bizony fel van adva a lecke, hogy mindez miképpen magyarázható. Az egyik (A) verzió szerint a petesejt egy abnormális extra osztódáson ment keresztül, és a két petesejtet két spermium termékenyítette meg, majd a fejlődő (vegyes, XX és XY sejtekből álló) embrió sejtjei adott pillanatban gondoltak egyet és, akárcsak a normális egypetéjű ikrek esetén, két csoportra szakadtak. A másik (B) elgondolás szerint, a "kettészakadást" két egészen különleges esemény előte meg: a két spermium teljesen egyszerre ért a petesejthez (ugyanis a megtermékenyített petesejt normális esetben azonnal blokkolja a fölösleges spermiumok behatolását), így egy triploid sejt keletkezett. Az első osztódás után keletkező két sejtbe azonban már csak normál, diploid genetikai állomány került, és az egyik sejtből az egyik spermium, a másikból a másik spermium kromoszómái vesződtek el.
Az egész eseménysorozat és jelenség szinte valószínűtlennek tűnik s valóban nem is tudunk más esetről. De mivel ezt is csak az egyik baba hermafroditizmusa okán szúrták ki, kérdés, hogy nem sokkal gyakoribb-e az ilyen fajta kimérizmus ikrek között, mint azt eddig gondoltuk?
Ross, CN, French, JA, Orti, G (2007) Germ-line chimerism and paternal care in marmosets (Callithrix kuhlii). PNAS doi: 10.1073/pnas.0607426104.
Souter, VL, Parisi, MA, Nyholt, DR, Kapur, RJ, Henders, AK, et al. (2007) A case of true hermaphroditism reveals an unusual mechanism of twinning. Hum Genet 121: 179-185.
 A tankönyvi történet szerint, az emlősök mai sokfélesége annak a radiációnak köszönhető, amely mintegy 65 millió évvel ezelőtt, a dinoszauruszok hirtelen kipusztulása után következett be. Egészen addig csak apró éjszakai állatokként húzódtak meg a dinoszauruszok árnyékában, s ha nincs a
A tankönyvi történet szerint, az emlősök mai sokfélesége annak a radiációnak köszönhető, amely mintegy 65 millió évvel ezelőtt, a dinoszauruszok hirtelen kipusztulása után következett be. Egészen addig csak apró éjszakai állatokként húzódtak meg a dinoszauruszok árnyékában, s ha nincs a  A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
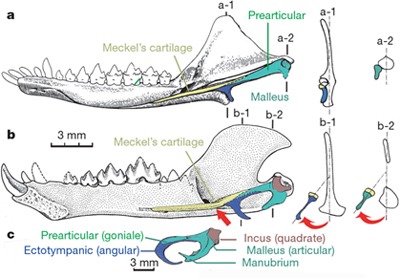 A nagy kérdés az tehát, hogy vajon mennyiben követhetjük az ominózous két csontocska átalakulását az emlősök evolúciója során. A válasz már eddig is az volt, hogy
A nagy kérdés az tehát, hogy vajon mennyiben követhetjük az ominózous két csontocska átalakulását az emlősök evolúciója során. A válasz már eddig is az volt, hogy 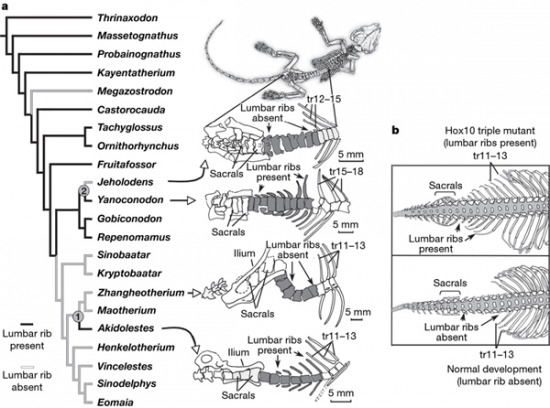
 A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő
A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő  A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.
A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.

{kind=link}
{kind=link}
{kind=link}
Utolsó kommentek